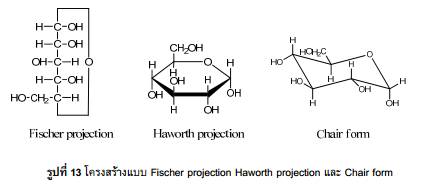
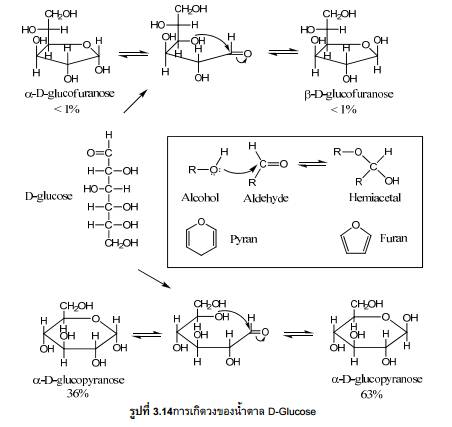
โปรตีน (Protein)
VIDEO
VIDEO
โปรตีนเป็นสารประเภทพอลิเพปไทด์ที มีโครงสร้างของโมเลกุลที ซับซ้อน มีธาตุประกอบ
หลักซึ งประกอบด้วย คาร์บอน ไฮโดรเจน ออกซิเจน และไนโตรเจน โปรตีนเป็นสารที พบมากที สุด
ในสิ งมีชีวิตมากกว่าครึ งหนึ งของนําหนักแห้ง (dry weight) ของเซลล์ทั่วไป โปรตีนมีบทบาทสําคัญ
ในกระบวนการทางชีวเคมีทุกชนิด ดังนั้นความรู้เกี่ยวกับโครงสร้างและการทํางานของโปรตีนจึงมี
ความสําคัญมาก
1. กรดอะมิโนและพันธะเพปไทด์ เมื อสลายโปรตีนด้วยกรดจะได้สารอินทรีย์ง่ายๆ เรียกว่า กรดอะมิโน (amino acid) ซึ งมี
สูตรโครงสร้างเป็น NH2
-CHR-COOH ในสูตรโครงสร้างนี้อะตอมคาร์บอนในตําแหน่งอัลฟาจะเป็น
อะตอมคาร์บอนไม่สมมาตร (asymmetric carbon atom) ดังนั้นกรดอะมิโนทุกตัว นอกจากไกลซีน
จะมีสเตริโอไอโซเมอร์ได้สองชนิด คือ D- และ L- โดยที กรดอะมิโนในธรรมชาติส่วนใหญ่เป็นชนิด L-isomer
โปรตีนเกิดจากกรดอะมิโนสร้างพันธะด้วยพันธะเพปไทด์ เชื่อมต่อกันระหว่างโมเลกุล ซึ่งมี
การจัดเรียงลําดับ ชนิด และจํานวนที แตกต่างกัน เกิดเป็นโครงสร้างโมเลกุลขนาดใหญ่ที มีมวล
โมเลกุลมาก
โปรตีนเป็นพอลิเมอร์ธรรมชาติที เกิดจากการรวมตัวของกรดอะมิโนจํานวนมาก โปรตีนมี
ตาราง โครงสร้างและชนิดของกรดอะมิโนที)มีโซ่ข้างมีขั้ว (Polar, Uncharged) (Garrett, 2548)
ตาราง โครงสร้างและชนิดของกรดอะมิโนทีมีโซ่ข้างเป็นกรด (Acidic) (Garrett, 2548)
ตาราง โครงสร้างและชนิดของกรดอะมิโนทีมีโซ่ข้างเป็นเบส (basic) (Garrett, 2548)
ในบางครั้งจําแนกสารตามความแตกต่างกันที โซ่แขนง หรือ R group ได้ 7 แบบได้แก่ กลุ่ม
aliphatic ได้แก่ ไกลซีน อะลานีน เวลีน ลูซีน และ ไอโซลูซีนกลุ่ม aromatic ได้แก่
เฟนิลอะลานีน ทริปโตเฟน และ ไทโรซีน กลุ่ม Sulphur ได้แก่ ซีสเตอีน และ เมไธโอนีน กลุ่ม
acidic ได้แก่ กรดแอสปาร์ติค และ กรดกลูตามิค กลุ่ม basic ได้แก่ ไลซีน อาร์จินีน และ
ฮีสติดีน กลุ่ม amide ได้แก่ แอสปาราจีน และ กลูตามีนกลุ่ม Hydroxy ได้แก่ เซรีน และ
ธรีโอนีน และกลุ่มสุดท้ายได้แก่ กลุ่ม Imino ได้แก่ โปรลีน
2. คุณสมบัติของกรดอะมิโน เนื่องจากกรดอะมิโนมีหมู่อะมิโนและหมู่คาร์บอกซิลิกเป็นหมู่ฟังก์ชันจึงมีสมบัติดังนี้
การละลายนํา กรดอะมิโนเป็นผลึกของแข็งสีขาว ละลายนําได้และละลายในโมเลกุลที มีขั้ว จุด
หลอมเหลวของกรดอะมิโนค่อนข้างสูงเมื่อเทียบกับสารอินทรีย์อื่นๆ ที่มีมวลโมลเลกุลใกล้เคียงกัน
เนื่องจากแรงยึดเหนี่ยวระหว่างโมเลกุลเป็นพันธะไฮโดรเจน ความเป็นกรด-เบสของกรดอะมิโนมี
สมบัติเป็นทั้งกรดและเบส เนื่องจากมีองค์ประกอบเป็นหมู่ฟังก์ชันของ –COOH ซึ งมีสมบัติเป็นกรด และหมู่ –NH2ซึ่งมีสมบัติเป็นเบส กรดอะมิโนจึงเป็นสาประเภทแอมโฟเทอริก ซึ่งแตกตัวเป็นไอออนในสารละลายได้คล้ายสารประกอบไอออนิก ต่างกันที่ โมเลกุลของกรดอะมิโนมีทั้งประจุบวกและประจุลบ จึงเกิดเป็นไอออน 2 ขั้วที เรียกว่า zwiiter ion ทําให้ผลรวมของประจุไฟฟ้าในโมเลกุลเป็นศูนย์
และเมื่อทําปฏิกิริยากับเบสจะได้
และเมื่อพิจารณาค่า pK ของกรดอะมิโนแต่ละชนิดตัวอย่างเช่น อะลานีนซึ งมีค่า pKaCเท่ากับ 2.4
และ pKaNเท่ากับ 9.9 หากนําค่า pKaC บวกกับ pKaNแล้วหาร 2 จะได้ค่า PKIซึ งเรียกค่านี0ว่า
Isoelectric point ซึ งเป็นค่าที ทําให้ประจุรวมของกรดอะมิโนมีประจุรวมเป็นศูนย์ แสดงดังสมการ
และเมื อพิจารณากรดอะมิโนที มีสมบัติเป็นกรด เช่นกรดแอสปาติคพบว่ามีค่า pKaCเท่ากับ 2.0pKaN
เท่ากับ 9.9 และ pKaRเท่ากับ 3.9 เมื อพิจารณาสมการเคมีพบว่าสามารถหาค่า pI ได้จาก ผลรวม
ของ pKaC กับ pKaR แล้วหาร 2 ได้ pI เท่ากับ 2.95
3. การสังเคราะห์ ααα-amino acids การสังเคราะห์กรดอะมิโนทําได้หลายวิธี วิธีการหนึ่งที ใช้ได้แก่วิธี Strecker Synthesis โดย
ใช้ ไฮโดรเจนไซยาไนด์และแอมโมเนียทําปฏิกิริยากับแอลดีไฮด์ ดังสมการ
4.โครงสร้างของโปรตีน
สิ่งมีชีวิตส่วนใหญ่ใช้กรดอะมิโนเป็นสารตั้งต้นในการสังเคราะห์โปรตีน โดยกรดอะมิโน
หลายโมเลกุลยึดเหนี ยวกันด้วยพันธะเพปไทด์ ถ้ากรดอะมิโน 2 โมเลกุลเกิดปฏิกิริยารวมตัวกันจะ
ได้สารประกอบที เรียกว่า ไดเพปไทด์ กรดอะมิโน 3 โมเลกุลทําปฏิกิริยารวมตัวกันจะได้
สารประกอบ ไตรเพปไทด์ และถ้ากรดอะมิโนหลายๆ โมเลกุลทําปฏิกิริยากันจนเกิดเป็นสายยาว
เรียกว่า พอลิเพปไทด์
ในการเขียนสูตรโครงสร้างเปปไทด์จะหันด้าน –NH2ไว้ด้านซ้ายมือ และ –COOH ไว้ด้านขวามือ
และเรียกด้านที มี -NH2ว่า N-terminal และ ด้าน –COOH ว่า C-terminal
5.การเรียกชื)อโปรตีน การเรียกชื่อโปรตีนไดเพปไทด์และไตรเพปไทด์ ให้เรียกชื อตามลําดับของกรดอะมิโน โดย
เรียกชื่อกรดอะมิโนลําดับแรกด้วยการลงท้ายด้วย –yl ตามด้วยชื อกรดอะมิโนตัวสุดท้าย
เช่น
6.การทดสอบพันธะเพปไทด์ การทดสอบพันธะเพปไทด์ด้วยสารละลายเจือจาง CuSO4ในสารละลายเบส เรียกว่าปฏิกิริยา ไบยูเรต ซึ่งจะให้ผลกับสารที ประกอบด้วยพันธะเพปไทด์ตั้งแต่ 2 พันธะขึ้นไป เป็นปฏิกิริยาที่ใช้ทดสอบสารประเภทโปรตีนได้ทุกชนิด รวมถึงผลิตภัณฑ์ที่เกิดจากการไฮโดรไลซ์โปรตีนที ยังเป็นโมเลกุลขนาดใหญ่ ซึ่งให้ผลการทดสอบเป็นสีนําเงินม่วง
7. ไฮโดรไลซิสเปปไทด
เมื่อโปรตีนหรือเปปไทด์ถูกreflux กับกรด HCl ความเข้มข้น 6M พันธะเอไมด์จะถูกย่อย
สลายกลายเป็นกรดอะมิโนได้ ซึ่งเราสามารถทดสอบว่ามีกรดอะมิโนหรือไม่โดยใช้ Ninhydrin โดย Ninhydrin จะทําปฏิกิริยากับ α-amino acids ให้สีม่วงเกิดขึ้น ซึ่ง Ninhydrin จะไม่สามารถ เกิดปฏิกิริยากับ proline ได้เนื องจาก proline มีตําแหน่ง α-amino acid เป็น secondary amide จึงไม่เกิดปฏิกิริยากับNinhydrin
8.โครงสร้างของโปรตีน
เนื่องจากโปรตีนเป็นโมเลกุลขนาดใหญ่ซึ่งประกอบด้วยกรดอะมิโนจํานวนมากซึ่งยึดเหนี่ยว
กันด้วยพันธะเพปไทด์และพันธะชนิดอื่นๆ ทําให้โปรตีนมีโครงสร้าง 4 ระดับคือ
8.1 โครงสร้างปฐมภูมิ (Primary structure)
เป็นโครงสร้างที แสดงการจัดลําดับ ชนิด และ จํานวนโมเลกุลของกรดอะมิโนในสายพอลิเมอร์โซ่ยาว ซึ่งโปรตีนแต่ละชนิดจะมลําดับของชนิดและจํานวนโมเลกุลของกรดอะมิโนที แน่นอน การจัดลําดับกรดอะมิโนโครงสร้างปฐมภูมิกําหนดให้ปลายหมู่อะมิโนอยู่ด้านซ้าย(N-terminal) และ ปลายคาร์บอกซิลิกอยู่ด้านขวา (C-terminal)
8.2 โครงสร้างทุติยภูมิ (secondary structure) เป็นโครงสร้างที เกิดจากการขดหรือม้วนตัวของโครงสร้างปฐมภูมิ ถ้าเกิดจากการสร้างพันธะไฮโดรเจนระหว่าง C=O ของ กรดอะมิโนหนึ งกับ N-H ของกรดอะมิโนถัดไปอีก 4 หน่วยในสายพอลิเพปไทด์เดียวกันจะเกิดโครงสร้างในลักษณะบิดเป็นเกลียวซึ งเรียกโครงสร้างทุติยภูมิชนิดนี้ว่า เกลียวแอลฟา และถ้าเกิดจากการสร้างพันธะไฮโดรเจนระหว่างC=O กับ N-H ของกรดอะมีโนระหว่างสายพอลิเพปไทด์ที่อยู่คู่กันจะเกิดโครงสร้างแบบแผ่น เรียกว่าแผ่นพลีทบีต้านอกจากจะเกิดพันธะไฮโดรเจนแล้วยังสามารถเกิดพันธะไดซัลไฟด์พันธะไอออนิก ฯ
8.3 โครงสร้างตติยภูมิ (tertiary structure)
ซึ่งประกอบด้วยโครงสร้างทุติยภูมิหลายส่วนรวมกัน โดยมีแรงยึดเหนียวอ่อนๆ คล้ายโครงสร้างทุติยภูมิ โครงสร้างตติยภูมิของโปรตีนแต่ละชนิดมีลักษณะจําเพาะขึ้นอยู่กับลําดับของกรดอะมิโนในสายพอลิเพปไทด์ ทําให้เกิดโครงสร้างที เหมาะสมในการทําหน้าที ต่างๆ ของโปรตีน
รูปที 3.4โครงสร้างทุติยภูม
8.4 โครงสร้างจตุรภูมิ (quaternary structure) เกิดจากการรวมตัวของ
หน่วยย่อยชนิดเดียวกันหรือต่างชนิดกันของโครงสร้างตติยภูมิ โดยมีแรงยึดเหนียวเหมือนกับใน
โครงสร้างทุติยภูมิและตติยภูมิลักษณะโครงสร้างใหม่ขึ้นอยู่กับโครงสร้างตติยภูมิซึ่งเป็นหน่วยย่อย
โดยอาจรวมกันเป็นลักษณะเป็นก้อนกลม เช่น ฮีโมโกลบินหรือเป็นมัดเส้นใย เช่น คอลลาเจน
9. ชนิดและหน้าที่ของโปรตีน การจัดจําแนกโปรตีนตามตามลักษณะโครงสร้างของสายพอลิเมอร์จําแนกได้ 2 ประเภาท
ดังนี้ โปรตีนเส้นใย (fiber protein) เกิดจากสายพอลิเพปไทด์หลายเส้นเรียงขนานกัน และพันรอบ
กันเองคล้ายเส้นเชือก ละลายนําได้น้อยส่วนใหญ่ทําหน้าที เป็นโปรตีนโครงสร้าง เพราะมีความ
แข็งแรงและยืดหยุ่นสูง ตัวอย่างโปรตีนชนิดนี้ได้แก่ ไฟโบรอินในเส้นไหม อีลาสตินในเอ็น คอลลา
เจนในเนื้อเยื่อเกี่ยวพัน เคราตินในผม ขน เล็บ เป็นต้น
โปรตีนก้อนกลม(globular protein) เกิดจากสายพอลิเพปไทด์ม้วนขดพันกันเป็นก้อนกลม
ละลายนํ0าได้ดี ส่วนใหญ่ทําหน้าที เกี ยวกับเมทาบอลิซึมต่างๆ ที เกิดขึ้นภายในเซลล์ ตัวอย่างของ
โปรตีนก้อนกลม เช่น เอนไซม์ ฮอร์โมนอินซูลิน ฮีโมโกลบิน โกลบูลินในพลาสมา เป็นต้น
นอกจากจะแบ่งโปรตีนตามโครงสร้างแล้วยังสามารถแบ่งตามหน้าที ได้ดังตาราง
ตาราง ชนิด หน้าที และแหล่งที พบของโปรตีน
9. เอนไซม์ (Enzyme) เอ็นไซม์เป็นโปรตีนที ทําหน้าที่ เป็นตัวเร่งปฏิกิริยาในเซลล์สิ งมีชีวิต ความหมายของเอนไซม์
คือ โปรตีนที มีลักษณะก้อนกลมทําหน้าที เร่งปฏิกิริยาการเปลี ยนแปลงของสารอาหารต่างๆ ใน
ร่างกาย และมีความจําเพาะต่อปฏิกิริยาและสารที ช่วยเร่งปฏิกิริยาโดยไม่ต้องเพิ มอุณหภูมิหรือ
เปลี ยนแปลงค่า pH ซึ งส่งผลให้ร่างกายสามารถควบคุมปฏิกิริยาต่างๆ ได้อย่างเหมาะสม
การเรียกชื อเอนไซม์ ให้เรียกชื อเหมือนสับสเตรทที เกิดปฏิกิริยากับเอนไซม์แล้วลงท้าย
พยางค์เป็น เ-ส เช่น ซูโครส เป็น ซูเครส อะไมโลส เป็น อะไมเลส มอลโทส เป็น มอลเทสเป็นต้น
9.1 สมบัติของเอนไซม์
1) มีความจําเพาะเจาะจง เอนไซม์ชนิดหนึ งใช้ได้กับสับสเตรตเพียงชนิดเดียว
เท่านั0น
2) เอมไซม์ทําหน้าที เป็นคะตะเลสที มีประสิทธิภาพสูง สามารถเร่งปฏิกิริยาได้หลาย
เท่ามากกว่าเมื อไม่ใส่เอมไซม์
9 .2 การทํางานของเอนไซม์ เริ่มต้นด้วยสารตั้งต้นที เรียกว่า สับสเตรต จับกับโมเลกุลของเอนไซม์ด้วยการต่อ
กับผิวของเอนไซม์ในส่วนที เรียกว่า บริเวณเร่ง (Active site) ซึ งเป็นบริเวณที มีความจําเพาะและ
สามารถต่อกันได้พอดีกับสับสเตรทเพียงชนิดเดียวเท่านั้น จึงเปรียบได้กับแม่กุญแจกับลูกกุญแจ
จากนั้นเอนไซม์กับสับสเตรตเกิดการเปลี่ยนแปลงไปเป็นสารประกอบเชิงซ้อน ซึ่งจะสลายตัวให้
ผลิตภัณฑ์และเอนไซม์กลับคืนมาซึ่งเอนไซม์เมื่อเกิดสารประกอบเชิงซ้อนจะทําให้พลังงานกระตุ้นมี
ค่าน้อยลงจึงทําให้เกิดปฏิกิริยาได้ง่ายขึ้น
ต่อมานักวิทยาศาสตร์พบว่าบริเวณตัวเร่งของเอนไซม์สามารถถูกเหนี ยวนําให้เหมาะสมกับ
สับสเตรตที เข้ามาจับหรืออาจเปลี ยนแปลงรูปร่างได้
9.3 ปัจจัยที)มีผลต่อการทํางานของเอนไซม์ 1) อุณภูมิ เอนไซม์จะทํางานได้ดีจะต้องอยู่ในอุณหภูมิที เหมาะสม
2) pHเป็นปจจัยหนึ งที มีผลต่อการทํางานของเอนไซม์ที ทําหน้าที เร่งปฏิกิริยา หาก
pH ไม่เหมาะสมจะทําให้เอนไซม์ทําหน้าที ไม่เต็มที่
3) ความเข้มข้นของเอนไซม์ หากเพิ มความเข้มข้นจะช่วยเร่งปฏิกิริยาให้เกิดเร็วขึ0น
แต่ถ้ามากเกินพออัตราการเกิดปฏิกิริยาจะมีค่าคงที
4) ปริมาณสารตั0งต้น มีผลต่ออัตราการเกิดปฏิกิริยาของเอนไซม์ เมื่อเพิ่มสับสเตรต
อัตราการเกิดปฏิกิริยาจะเพิ่มขึ้น แต่เมื่อยิ่งเพิ่มปริมาณของสับสเตรตมากเกินพอ อัตราการ
เกิดปฏิกิริยาจะไม่เกิดเร็วขึ้น เมื่อระดับหนึ่งอัตราการเกิดปฏิกิริยาจะคงที่ เนื่องจากไม่ได้เพิ่มปริมาณ
ของเอนไซม์
10. การแปลงสภาพโปรตีน โปรตีนแต่ละชนิดมีโครงสร้างสามมิติที จําเพาะต่อการทํางานที แตกต่างกัน ถ้าแรงยึดเหนี ยว
ในโครงสร้างสามมิติของโปรตีนถูกทําลาย จะทําให้โครงสร้างของโปรตีนเปลี ยนแปลงไป โดยการ
คลายตัวออกและไม่สามารถทําหน้าที ทางชีวภาพได้ เรียกว่า การแปลงสภาพโปรตีน ซึ งปจจัยที ทําั
ให้โปรตีนเกิดเปลี ยนสภาพได้แก่ ความร้อน pH เติมเอทานอลการเติม Pb(NO3)2ลักษณะการแปลงสภาพของโปรตีน ได้แก่ การแข็งตัว ไม่ละลายนํา เกิดตะกอนขุ่นขาว จับตัวเป็นก้อนขุ่นขาว เนื่องจากเสียสภาพทางชีวภาพและความเป็นระเบียบของโครงสร้าง กลไกการเสียสภาพโปรตีนใน
สภาวะที เป็นกรด-เบส ได้แก่ โปรตีนจะให้หรือรับ H+ แล้วเกิดเป็นไอออน สามารถจับไอออนอื่นได้
และไอออนลบที แตกตัวออกจากสารละลายกรดเป็นไอออนขนาดใหญ่ จึงทําให้ไอออนของโปรตีน
รวมตัวกันเป็นก้อนได้ ในตัวทําละลายอินทรีย์สารจะเกิดพันธะไฮโดรเจนกับโปรตีน ทําให้โปรตีน
แข็งตัวและละลายนําได้น้อยลง ในสารละลายที มีไอออนของโลหะหนัก เช่น Pb2+ และ Ag+
จะเกิดการจับตัวระหว่างไอออนบอกของโลหะหนักกับกรดอะมิโนในโปรตีนด้านคาร์บอกซิล เกิดเกลือคาร์บอกซิเลต ซึ งละลายนําได้น้อยลง จึงเกิดการตกตะก้อน